基因科普:系统解读基因编辑领域的主要技术
在生命科学领域,人们一直有一个目标,即能够程序化、特异性和高效地编辑所有活细胞的DNA序列,这在基因研究、基因治疗、遗传育种和合成生物学中具有无限的价值。
以前的一些方法,如巨核酸酶、锌指核酸酶(ZFN)和转录激活因子样效应核酸酶(TALEN),依赖于复杂的特定的蛋白质-DNA相互作用,将蛋白质效应物靶向所需的DNA序列。虽然对于靶向特定位点有效,但很难简单迅速地将这些蛋白质域的目标重新编程为感兴趣的新基因组位点。后来, CRISPR系统的发现和工程化应用在基因组编辑领域引发了一场新的令人兴奋的革命。
近日,张先恩教授、刘陈立研究员、戴俊彪研究员、元英进教授、高彩霞研究员、冯雁教授、吴边研究员、魏平研究员、游淳研究员、汪小我副教授、司同研究员在
Science China Life Sciences 期刊上发表了题为:Enabling technology and core theory of synthetic biology 的综述论文,详细论述了合成生物学领域的核心理论与技术。
本文编译了其中基因编辑技术的内容,供大家参考。
严格基于蛋白质的基因组编辑系统
巨核酸酶、ZFNs和TALENs是可用于基因组编辑的强大生物工具。
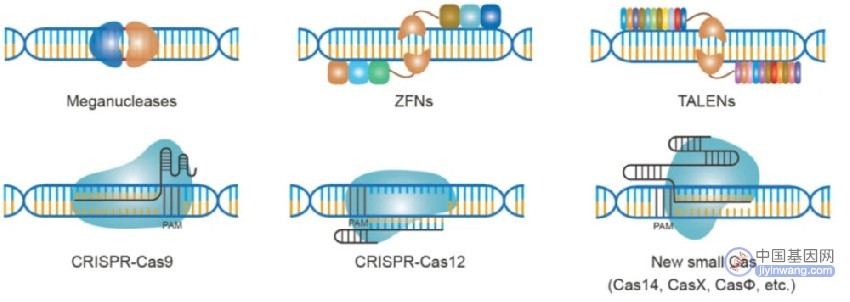
基于核酸酶的靶向DNA的基因组编辑技术,包括巨核酸酶,ZFNs,TALENs,CRISPR-Cas9,CRISPR-Cas12和新的小Cas变体
巨核酸酶(又称归巢核酸内切酶)是识别特定DNA序列的大型蛋白质复合体。这些蛋白质依赖于蛋白质本身和目标DNA序列之间复杂的相互作用网络。尽管之前的研究已经成功将巨核酸酶应用于新的、用户定义的基因组序列,但这一过程极其费力、耗时且技术上具有挑战性。巨核酸酶的应用从根本上依赖于整个蛋白质复合体的大规模重新编程,使它们能够识别新的感兴趣的DNA区域。因此,迫切需要使用需要变异库的高通量方法来识别新定义的蛋白质序列识别新的变异体。因此,需要更多的可编程方法来实现热稳定性DNA的靶向。
锌指蛋白是能够识别三种DNA碱基特定序列的小蛋白质模块。这些蛋白质在自然界很常见,以前的研究已经确定了单个锌指模块的关键成分,这些模块决定了特定的3碱基对DNA结合序列。可以将模块化的锌指阵列融合在一起,以实现基于特定DNA序列的DNA靶向。此外,研究人员巧妙地将这些参与DNA靶向的较大锌指蛋白与可以切割DNA的FokI蛋白融合在一起。为了最大限度地减少活细胞中所有不需要的随机DNA切割,研究人员巧妙地将FokI蛋白分成两部分,每一半使用特定的锌指招募到DNA的目标区域。因此,两个锌指核酸酶(ZFNs)的结合可以特异性地、准确地切割DNA。这些ZFNs已被证明在人类、动物和植物细胞中发挥功能,因此在可编程基因组编辑中发挥重要作用。
随着锌指蛋白的发现,研究人员从植物病原体中鉴定出转录激活物样(TAL)效应器。与ZFNs不同,每个TAL效应器(TALE)与单个DNA碱基结合。这种效应可以通过编程与特定的DNA序列结合。TALEs与FokI二聚体结合以产生TALE核酸酶(TALENs),这是一种完全基于蛋白质的可编程基因组编辑技术。与ZFNs相比,TALENs显示出更好的编程能力,因为每个DNA碱基都由一个单元识别,而不是锌指的三重编码特性,但TALENs比ZFNs大,因此其递送问题仍然具有挑战性。此外,需要构建蛋白质复合体,这在寻求广泛编辑活细胞基因组时并不容易。
CRISPR-Cas系统
在研究细菌基因组时,研究人员发现了一段重复的DNA,命名为规律成簇的间隔短回文重复序列(CRISPR)。通过随后的研究,研究人员证明了CRISPR序列及其附近的蛋白质,即CRISPR相关(Cas)蛋白,可以作为细菌免疫系统来抵御外来入侵的核酸。当细菌暴露于致病DNA片段时,免疫系统分离一小部分外源DNA,并将该序列整合到细菌基因组本身的CRISPR序列中。这一发现对于CRISPR-CAS这一项革命性的基因组编辑技术的发展至关重要。
CRISPR阵列被鉴定为编码与Cas蛋白相关的RNA序列,并以DNA中基于Protspacer(原间隔序列)的核酸序列为靶标。随后的工程证明,CRISPR RNA中的靶向序列可以很容易地用用户定义的序列替换和编程,这将完全改变和重新编程Cas蛋白靶向序列。这一发现在基因组编辑领域发挥着重要作用,因为这是第一次通过替换核酸序列来轻松地实现基因组编辑,这与以前需要复杂的高通量的蛋白质工程的方法不同。一旦与目标DNA序列结合,Cas蛋白就会启动双链DNA的切割,对活细胞的基因组造成损伤。
新型CRISPR蛋白
Cas蛋白是CRISPR基因组编辑技术的关键组成部分。来自化脓性链球菌的Cas9(spCas9)是第一个用于基因组编辑应用的工程化Cas蛋白,在开发新的编辑技术时将继续受到广泛使用。已知所有Cas蛋白都需要原型间隔子相邻基序(PAM),这是一小段直接位于目标基因组位点附近的DNA。当我们试图在细胞基因组的其他位置进行编辑时,Cas蛋白的这种靶向范围限制将是一个挑战。研究人员发现了大量具有不同PAM要求的新型Cas蛋白,从而扩大了CRISPR基因组编辑技术的靶向范围。此外,蛋白质工程和定向进化方面的成就已成功改变Cas蛋白的PAM需求,为使用CRISPR/Cas系统开发一系列基因组靶标做出了贡献。
最近,许多新的小型Cas蛋白被发现。SpCas9的长度为1368个氨基酸,通过碱基编辑器和先导编辑器中的效应蛋白进一步延长。基因组编辑蛋白的稳定性和传递受到蛋白长度增加的负面影响。新的CRISPR-Cas蛋白,例如Cas12f,Cas14、CasΦ、CasX,比许多以前发现的Cas蛋白更小。然而,还需要进一步的工程优化、实验发现和技术进步来提高这些新Cas蛋白的编辑效率。
基因敲除
归巢核酸内切酶,ZFNs,TALENs和DNA靶向CRISPR-Cas系统都通过切割双链DNA发挥作用。在DNA双链断裂(DSB)产生后,细胞的内源性修复机制迅速修复病变。完美修复可以作为额外编辑的底物,直到非同源末端连接(NHEJ)或微同源介导的末端连接(MMEJ)修复在靶位周围产生随机的小DNA插入或缺失(INDEL)。INDEL导致基因敲除,这在某些情况下很有用,但缺乏精确性。同源定向修复(HDR)是一种竞争性修复过程,使用核酸供体模板来修复DNA。虽然可编程,但与NHEJ/MMEJ修复相比,HDR效率极低。因此,需要新的基因组编辑技术来高效、精确地编辑DNA序列。
碱基编辑
碱基编辑是一种可编程、高效且具有精确性的基因组编辑技术,其技术基础是将DNA结合蛋白定位到感兴趣的序列。设计的第一类碱基编辑器称为胞嘧啶碱基编辑器(CBE),利用了Cas蛋白结合DNA目标区域并将其解旋成单链DNA状态的能力。
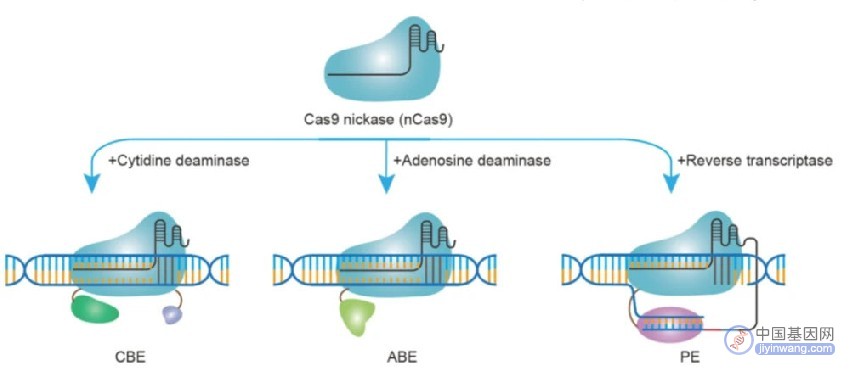
精准DNA基因组编辑技术包括包括胞嘧啶碱基编辑器(CBE)、腺嘌呤碱基编辑器(ABE)和先导编辑器(PE)
CBE由一种融合到Cas蛋白的单链特异性胞苷脱氨酶组成,可使Cas蛋白靶向的内源DNA区域脱氨基。DNA中胞嘧啶碱基脱去氨基产生尿嘧啶,尿嘧啶可通过内源性细胞过程复制并修复为胸腺嘧啶。为了提高编辑效率,CBE还加入了尿嘧啶糖基化酶抑制剂(UGI)来抑制内源性尿嘧啶N-糖基化酶(UNG),UNG可以特异性识别细胞基因组中尿嘧啶碱基的存在。局部UGI的存在将进一步延长尿嘧啶中间体的寿命,从而促进修复后胸腺嘧啶的永久结合。
为了进一步促进编辑,将Cas蛋白转化为切割酶,利用含有新的碱基编辑尿嘧啶的DNA的相反侧链作为修复模板,切割与编辑过的链相反的链,操纵细胞修复机制来替换被切割的微量链。最终实现从一条DNA链到两条DNA链的永久编辑,显著提高碱基编辑的效率。
腺嘌呤碱基编辑器(ABE)是开发的第二类碱基编辑器。ABE由实验室进化的腺苷脱氨酶组成,可将DNA中的腺嘌呤碱基转化为肌苷。肌苷随后被内源性细胞聚合酶识别为鸟嘌呤。先进的定向进化方法进一步提高了编辑效率并扩大了腺嘌呤碱基编辑的实用性。
最初的碱基编辑方法使用Cas蛋白来解开DNA,并暴露单链DNA序列作为脱氨的底物。一类名为DddA衍生胞嘧啶碱基编辑器(DdCBEs)的新型碱基编辑器利用一种称为DddA的天然双链DNA胞苷脱氨酶在不解开DNA的情况下进行碱基编辑。DNA结合蛋白,如TALEs或ZFs,可以融合分裂DddAs和UGIs,在没有Cas蛋白的情况下指导胞嘧啶碱基编辑靶向DNA序列。此外,通过将催化受损的DddA变异体与腺嘌呤脱氨酶TadA8e 融合,在人类线粒体中实现了靶向A到G的编辑。
CBE、ABE和DdCBE 都可以精确高效地编辑DNA,以创建CG到TA(CBE和DdCBE)或AT到GC碱基(ABE)的转换。然而,还有许多其他类型的基因组编辑,例如其他碱基转换以及可编程插入和删除,这些都需要更新的精确编辑技术。
先导编辑
先导编辑是一种精确的基因组编辑技术,这项技术利用了Cas蛋白结合DNA并切开一条DNA链的能力。与碱基编辑器相反,先导编辑器在Cas蛋白结合后切割未解开的单链R-环DNA。在对这条链进行特定的切割后,释放的DNA可以作为引物进行后续的DNA聚合。
先导编辑器的另一个关键组成部分是pegRNA,它编码一个引物结合位点,该位点与Cas蛋白缺口释放的单链R-环和编码特定所需DNA编辑事件的模板区域互补。RNA-DNA杂交后,与Cas蛋白融合的逆转录酶可以使用pegRNA作为模板来扩展基因组DNA。靶向先导编辑3'区域的正交Cas蛋白引导RNA可以用于进一步增强编辑。在随后的DNA复制和修复之后,新合成的DNA序列可以永久地整合到基因组中,从而产生由pegRNA序列指示的可编程和多功能编辑。先导编辑的初始演示在编辑效率方面相对较低,然而,随后对该程序进行了修改,例如优化引物结合熔化温度,使用两种pe-gRNA,DNA修复操作, RNA稳定性基序,以及逆转录酶修饰,大大提高了启动编辑效率。
RNA编辑
RNA中的基因组编辑可以避免对基因组的永久性改变,从而降低脱靶DNA编辑的风险。一类靶向Cas蛋白的RNA,如Cas13a和Cas7-11可编程地靶向由CRISPR引导序列确定的RNA序列。
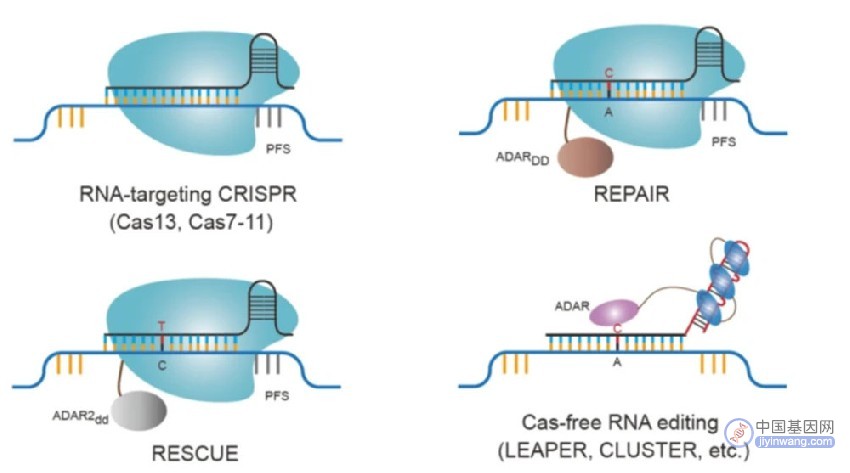
RNA编辑技术,包括CRISPR-Cas13,CRISPR-Cas7-11和RNA碱基编辑方法,如REPAIR,RESCUE和其他无CasRNA编辑方法
与DNA碱基编辑器的开发类似,研究人员通过将RNA特异性腺苷脱氨酶融合到RNA靶向Cas蛋白中来开发RNA碱基编辑器。腺苷脱氨酶RNA特异性(ADAR)蛋白通过“可编程A-to-I替换(修复)的RNA编辑”技术与Cas13a融合,将腺嘌呤转化为肌苷(类似于ABE DNA碱基)。同样,可以使RNA中的胞嘧啶脱氨的工程ADAR蛋白用于开发“特定CU交换的RNA编辑(RESCUE)”技术,该技术能够将胞嘧啶碱基转化为RNA中的尿嘧啶(类似于CBE DNA碱基编辑器)。
新的CRISPR-free RNA编辑系统已经开发问世,其能够利用RNA核酸招募内源性蛋白质与RNA发生化学反应的能力来进行位点特异性RNA编辑。此外,一项并行技术表明,较长的RNA可以自然地招募ADAR用于RNA的A-to-I编辑。最近,RNA的聚合设计和环设计大大提高了RNA编辑技术的编辑效率和特异性。
过去十年的特点是新的基因组编辑技术的快速发展。从最初的基于蛋白质的方法到精确的基因组编辑技术,如先导编辑,操纵活细胞和生物体基因组的能力越来越令人喜悦。RNA编辑技术也开始变得更加精确和高效。迫切需要继续开发更小、更精确、准确且高效的基因组编辑工具,特别是在治疗学、农业和生物研究等领域的应用方面。
基因组编辑的应用
基因组编辑技术的发展使整个生物医学和农业都取得了巨大的进步。生物技术公司在生产新的基因组编辑药物方面取得了一系列进展。最近,研究人员推进了体内基因组编辑技术的发展,例如CRISPR-Cas9和碱基编辑,以治疗镰状细胞性贫血、早老性痴呆、转甲状腺素淀粉样变或高胆固醇血症等遗传疾病。
基因组编辑在农业中的应用引发了对未来生物作物育种的全新憧憬。抗病和抗除草剂是许多作物品种中最发达的两个品种。最近,研究人员证明,四个同时发生的多重编辑事件能够提高作物抗病性并提高小麦植株的产量。此外,许多内源编辑已被证明能产生强大的除草剂抗性。基因组编辑将使持续创造有价值的农作物物种成为可能。
声明:本站所有文章资源内容,如无特殊说明或标注,均为采集网络资源。如若本站内容侵犯了原著者的合法权益,可联系本站删除。



















?")

