破解20年难题!《Nature》重磅:揭示调控细胞内基因转录的“红绿灯”机制!
假如基因是一条条设定好的马路,那么基因的转录过程就像是车辆在路上行驶,路况瞬息万变。未改变DNA序列的表观遗传修饰则如同道路上附属的信号灯、指示牌、加油站,调控着车辆的运行。如若信号灯出现了故障,交通就容易失控;如若基因的表观遗传修饰失调,基因的转录则会变得紊乱,进而引发癌症等疾病。
在诸多表观遗传的修饰方式中,组蛋白的甲基化修饰与基因的转录调控息息相关,其中组蛋白H3赖氨酸K4被广泛研究。三甲基化的组蛋白H3赖氨酸K4(H3K4 me3)对正常细胞发育至关重要,且与白血病、乳腺癌、肠癌和胰腺癌联系紧密。因此,了解它如何在正常细胞中发挥作用也可以为癌症的发展提供新的思路。但直到现在,科学家们仍对它的作用缺乏了解。
3月1日发表在《Nature》杂志上的一项重要新研究就揭示了H3K4me3控制细胞内基因转录的“红绿灯”机制【1】。在这项研究中,伦敦癌症研究所(The Institute of Cancer Research, London)的科学家们利用小鼠干细胞和复杂的遗传和生化实验,发现三甲基化的组蛋白H3赖氨酸K4(H3K4 me3)能够确保基因在正确的时间以可控的方式被转录和激活,就像一组信号灯在繁忙的道路上调节汽车的流量。

通过调节 RNA 聚合酶 II(一种读取和解码 DNA 的蛋白质复合物)的流动,H3K4me3 决定基因表达何时开始以及它运行的速度。当信号灯处在“绿灯”状态时,H3K4me3 允许 RNA 聚合酶 II 沿着 DNA 移动并将其转录成 RNA。但若没有 H3K4me3,RNA 聚合酶 II 会卡在 DNA 的启动子近端区域,造成阻碍减慢转录。
建立研究H3K4甲基化的模型
H3K4的甲基化受SET1/COMPASS复合物的直接调控。SET1/COMPASS由多个赖氨酸甲基转移酶和若干个亚单位构成,因此各个蛋白构建存在功能冗余。而这也使确定 H3K4 甲基化在基因表达中的直接作用变得十分困难。为研究H3K4 甲基化在转录中的作用,研究人员开发了DPY30 或 RBBP5 的急性耗竭(acute depletion)模型系统。之所以选择DPY30 和 RBBP5是因为这两个核心成分是小鼠胚胎干 (mES) 细胞系中所有 SET1/COMPASS 复合物家族不可或缺的共享成分。为了构建急性耗竭模型,研究人员在mES 细胞中表达了DPY30 或RBBP5与degron(一种可促进蛋白降解的蛋白)的融合蛋白。其中DPY30与受生长素调控的mAID degron tag融合,RBBP5则与受dTAG-13调控的FKBP degron tag融合。用该细胞模型进行实验的结果表明,DPY30 或 RBBP5 的急性耗竭会导致 H3K4 甲基化的快速丧失。
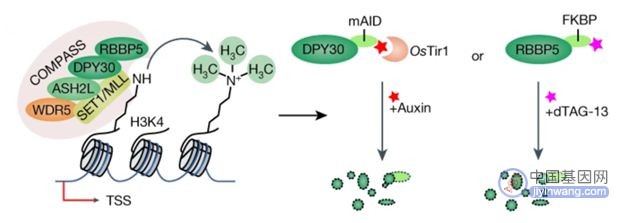
图注:靶向DPY30 或 RBBP5的 degron系统【1】
去甲基化酶KDM5 调控
H3K4me3 的快速转化
那么,H3K4me3 的快速丧失是受谁的调控呢?组蛋白转化和主动去甲基化是2个可能的机制。为了研究 H3K4me3 的快速转化是否是由于 KDM5 H3K4me3/me2 去甲基化酶的活性,研究人员敲除了 DPY30–mAID mES 细胞系中的Kdm5a和Kdm5b基因。他们发现与 DPY30-mAID 细胞相比, H3K4me3 在 Kdm5a/b敲除细胞(dKO)中持续存在时间显著延长,表明H3K4me3 的快速转化依赖于 mES 细胞中的 去甲基化酶KDM5A 和 KDM5B。
基因转录的调控需要H3K4me3
为了研究H3K4me3 快速丧失后对转录的主要影响,研究人员使用 SLAM-seq 在 degron 细胞系中测量了新转录 RNA 的合成速率。用生长素处理 dKO 细胞和DPY30-mAID 细胞后进行的 SLAM-seq 分析发现,DPY30-mAID细胞下调的基因数量远多于dKO细胞,说明H3K4me3 是调节转录的决定性因素。
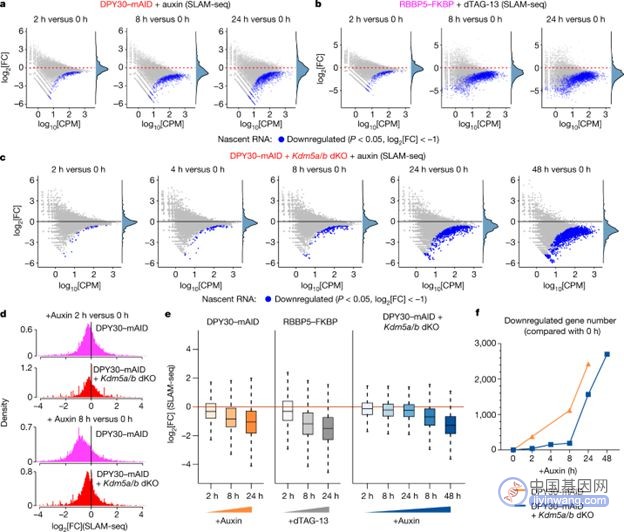
图注:基因转录的调控需要H3K4me3【1】
H3K4me3是如何调控转录的呢?研究人员发现H3K4me3 的快速丧失会促进RNA 聚合酶II 在启动子区域的暂停 (RNAPII pausing)。他们进一步利用高分辨率的mNET-seq对dKO细胞和DPY30-mAID进行分析,发现与DPY30-mAID细胞相比,dKO细胞的RNAPII pausing存在显著延迟,表明H3K4me3参与调控RNA 聚合酶II的暂停与释放。
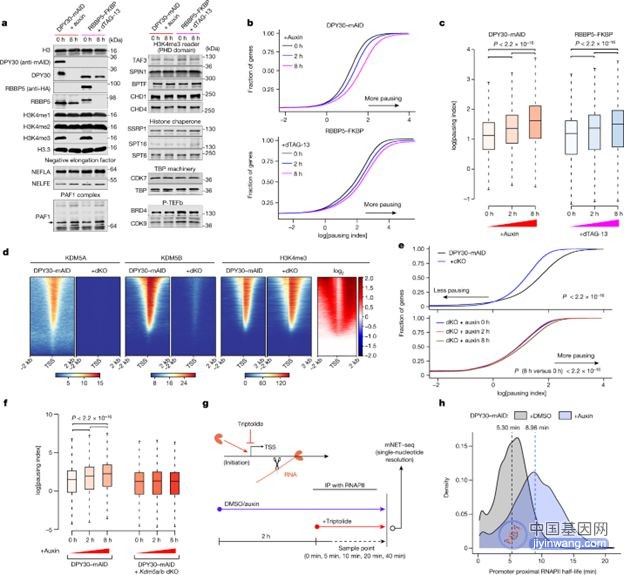
图注:H3K4me3 的快速丧失延长 RNAPII pausing的停留时间【1】
那么,H3K4me3通过RNA 聚合酶II的调控对基因转录产生了怎样的影响呢?研究人员利用TTchem-seq和mNET-seq技术测量的转录延伸速率,发现H3K4me3快速丧失会降低延伸速率。
INTS11参与H3K4me3调控的启动子近端暂停-释放
(promotor proximal pausing-release)
研究人员接下来探究可能参与受H3K4me3调控的启动子近端暂停释放的蛋白质。他们利用ChIP-MS识别了3个蛋白并对其中的INTS11(一个核酸内切酶亚基)做后续研究。他们发现与H3K4me3相似,INTS11的快速丧失会促进RNA 聚合酶II的暂停。进一步显示H3K4me3 通过招募INTS11 的机制调节启动子近端暂停,这于RNAPII pausing的取消和转录延伸至关重要。
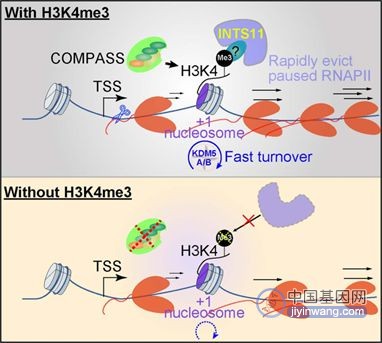
图注:H3K4me3调控转录的机制【1】
此项研究的负责人,表观遗传学研究专家、伦敦癌症研究所的首席执行官Kristian Helin教授表示:“我们的研究提供了对表观遗传学的新理解,这是一个非常令人兴奋但仍未充分探索的癌症领域研究。揭示了一种著名的表观遗传修饰如何控制基因表达,解决了一个存在 20 年的难题。由于决定细胞中 H3K4me3 水平的酶经常在癌症中发生突变,这项研究可能对理解和治疗癌症具有重要意义。”【2】
参考文献:
【1】Wang, H., Fan, Z., Shliaha, P.V. et al. H3K4me3 regulates RNA polymerase II promoter-proximal pause-release. Nature (2023). https://doi.org/10.1038/s41586-023-05780-8
【2】New study unveils epigenetic 'traffic lights' controlling stop and go for gene activity (2023, March 1) retrieved 3 March 2023 from https://medicalxpress.com/news/2023-03-unveilsepigenetic-traffic-gene.html
声明:本站所有文章资源内容,如无特殊说明或标注,均为采集网络资源。如若本站内容侵犯了原著者的合法权益,可联系本站删除。





")















