真核生物的基因表达调控,即从DNA到蛋白质的调节过程
1、真核生物调控蛋白
细菌的调控蛋白大多数利用螺旋-转-螺旋基序结合 DNA,大多数以二聚体的形式结合DNA,一个螺旋(识别螺旋)插入 DNA 大沟相契合,识别特殊的碱基对;另一个螺旋与DNA骨架接触,使识别螺旋正确定位,并增强结合;
真核生物调控蛋白细节上有差异,识别 DNA 的原理与细菌类似,除了同源二聚体,一些调控蛋白形成异源二聚体识别DNA,增加了可以特异结合的DNA的范围;真核细胞活化子的 DNA 结合和激活功能是分开的;激活域没有确定结构,根据氨基酸组成可分为酸性激活域;谷氨酰胺富集区;脯氨酸富集区;
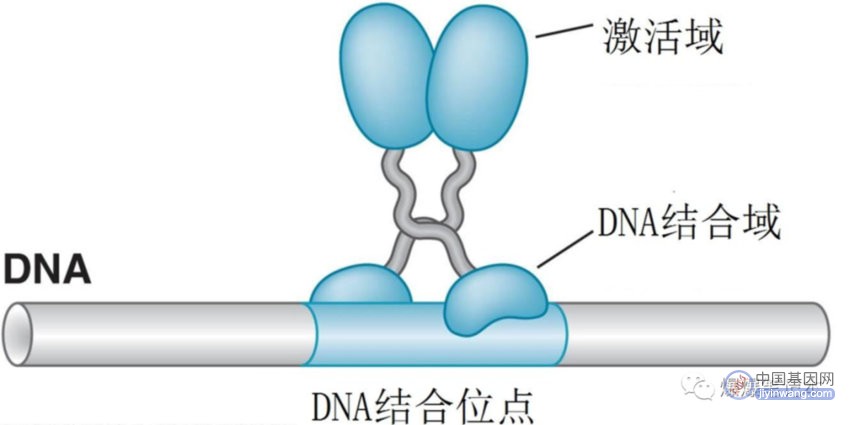
1.1 真核细胞调控蛋白的DNA结合域
真核细胞调控蛋白识别DNA的原理与细菌的相同,有多种DNA结合域;
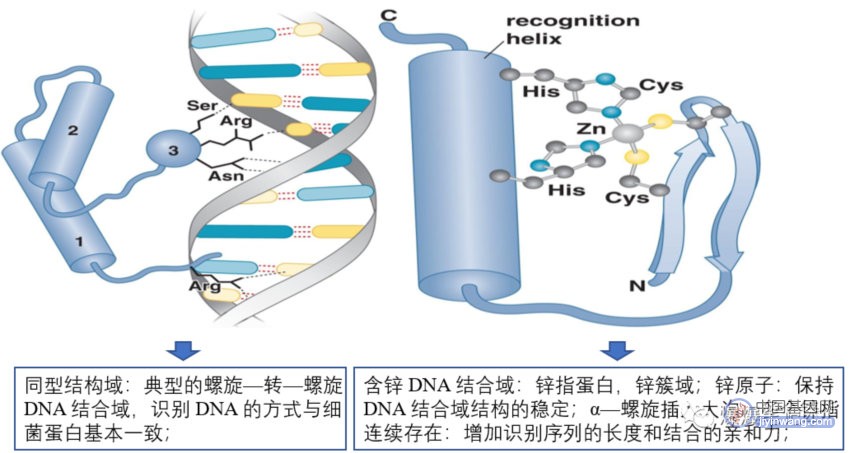
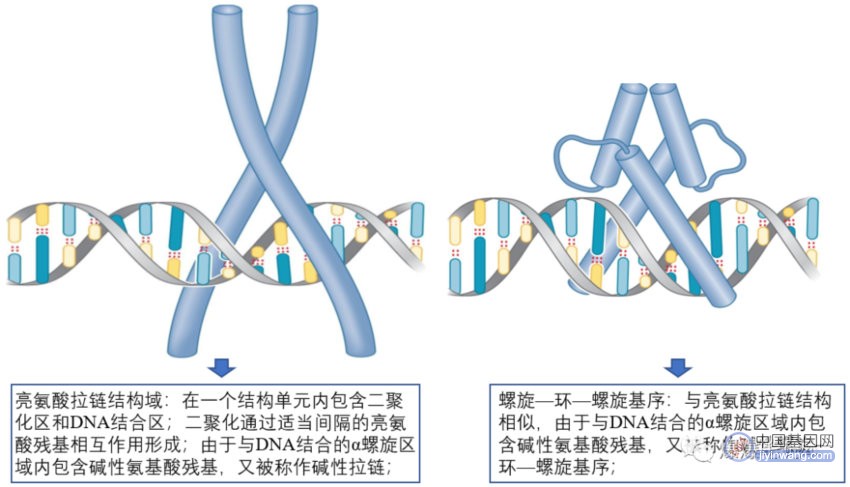
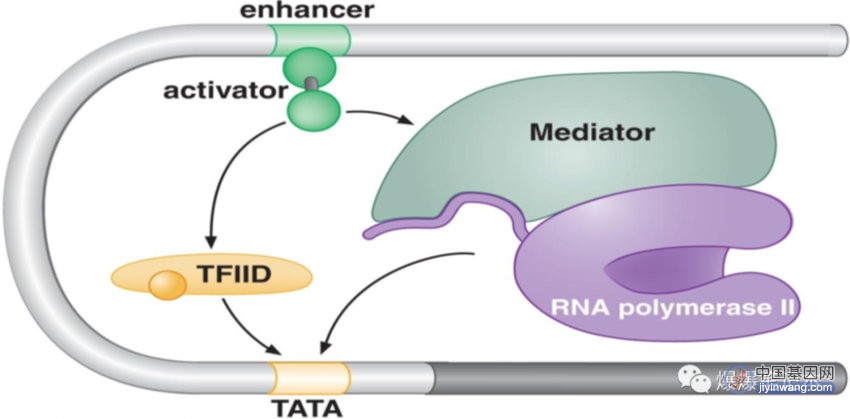
1.2 活化子与真核转录机器蛋白的协同作用
除了 RNA 聚合酶,真核转录机器还包括很多其他蛋白质. 这些蛋白质形成几个复合物,如中介蛋白和 TFIID 复合物. 活化子与一个或多个复合物相互作用,将它们招募到基因上. 其他没有被活化子直接招募的成分,通过已被招募成分协同结合.
1.3 活化子募集核小体修饰物
活化子也募集核小体修饰物帮助转录机器结合到启动子上或帮助转录起始;募集的核小体修饰物有两类:一类是在组蛋白尾巴上加化学基团,如乙酰基团(是由组蛋白乙酰转移酶进行的);第二类是可以重塑核小体的重塑复合物,如依赖ATP活性的SWI/SNF蛋白复合物;核小体修饰物通过核小体重塑和组蛋白修饰,如乙酰化,使DNA松散,暴露核小体内部的原本无法接近的DNA结合位点,激活基因表达;下图为活化子指导的染色质局部结构改变:
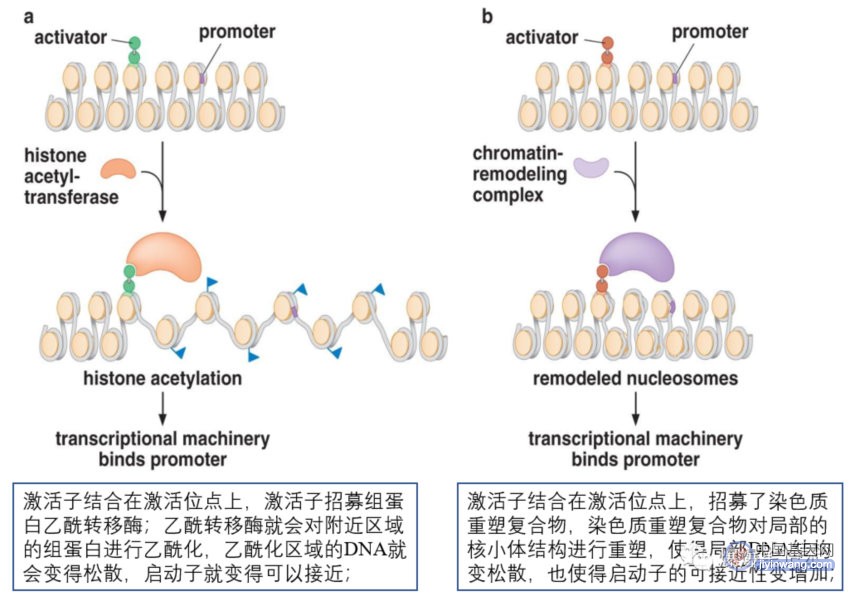
乙酰基团还以另一种方式促进转录机器的结合:TFHID复合物的一个组分含有特异性结合乙酰基团的结构域—bromodomain,若某个基因的启动子含有乙酰化的核小体,就会对转录机器有更高的亲和力;
1.4 多活化子的协同作用
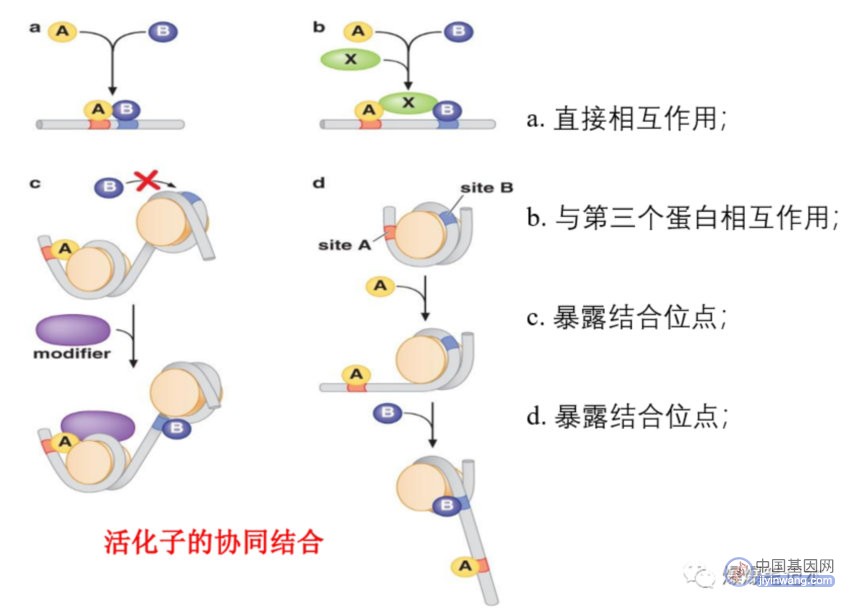
多细胞生物中信号整合更广泛存在,某些基因的表达需要多个信号,每个信号通过独立的调控蛋白传给基因,因此,多个活化子常共同作用,且常常协同作用(两种活化子共同作用产生的效应大于二者分别作用所产生效应的简单加和);协同作用的三种策略:1. 多个活化子招募转录机器的同一成分,如:与中介蛋白不同部位的接触,组合结合的能量对招募有指数效果;2. 多个活化子分别招募转录机器的不同组分,若没有帮助都不能有效结合启动子;3. 多个活化子相互帮助能更好地与所调控基因上游的位点相结合;
2、基因沉默
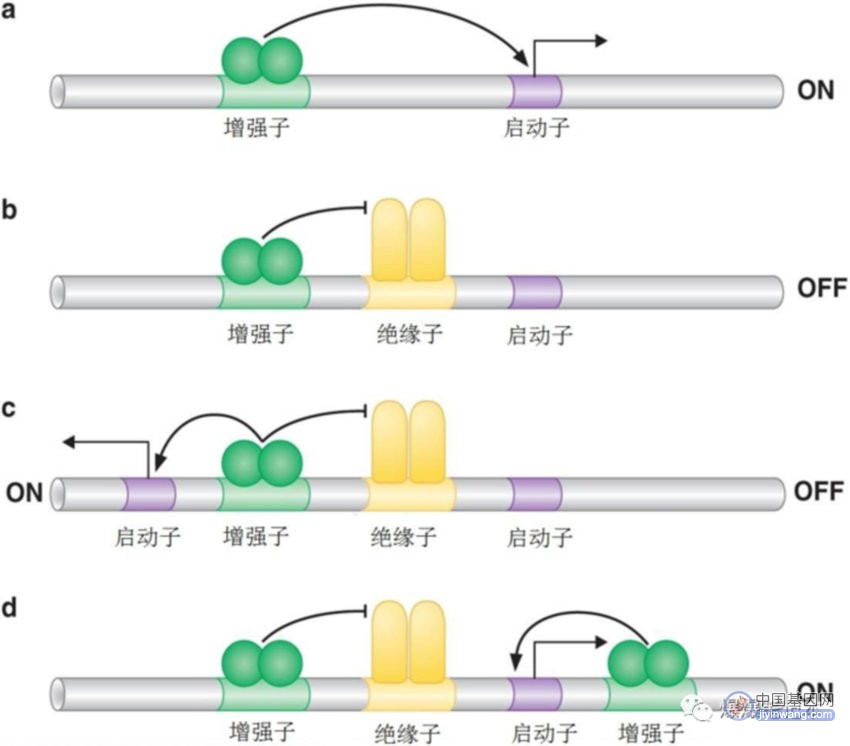
基因“沉默”是一种位置效应,基因由于它所处的位置而沉默,不是对特定环境信号的反应,沉默效应可在大段DNA序列上扩散,“沉默”多个基因,不需要每个基因都有特定抑制子结合位点;包括离起始事件距离较远的基因;绝缘子元件能够阻断沉默的扩散;绝缘子能使基因免于不加选择的活化和抑制;(绝缘子的应用:实验中,随机插入哺乳动物基因组的基因经常处于沉默状态(插入到异染色质区);在该基因的上游和下游加入绝缘子可使该基因将免于沉默).最常见的沉默与异染色质有关,异染色质通常与染色体的特殊区域相关,如端粒和着丝粒,哺乳动物细胞中约50%的基因组以异染色质形式存在;转录沉默常与核小体修饰有关,改变基因对转录机器和其他调控蛋白的可接近性,DNA甲基化也可沉默转录(DNA甲基化是由DNA甲基化酶完成的);绝缘子元件能够阻断沉默的扩散;绝缘子能使基因免于不加选择的活化和抑制;(绝缘子的应用:实验中,随机插入哺乳动物基因组的基因经常处于沉默状态(插入到异染色质区);在该基因的上游和下游加入绝缘子可使该基因将免于沉默).
顺式作用元件绝缘子控制活化子,阻止非特异性基因激活:
2.1 酿酒酵母中的基因沉默
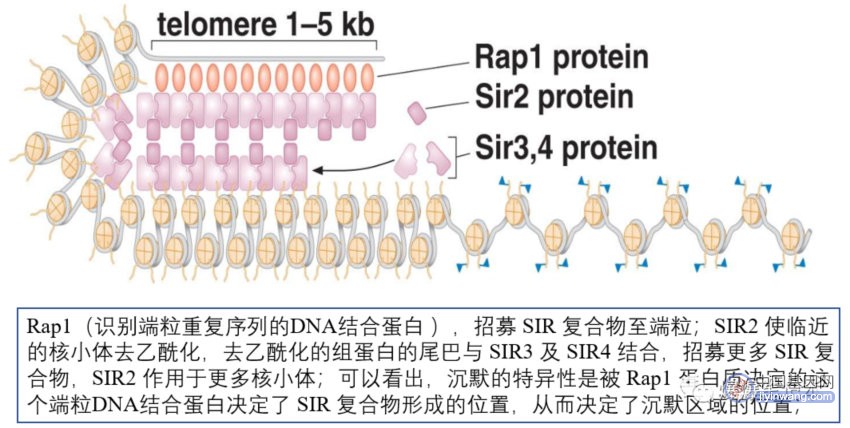
在酿酒酵母中的“沉默”区域包括端粒、沉默的交配型基因座和 rDNA;以端粒为例:染色体末端1-5 kb,紧密结构,乙酰化程度低,插入此区域的基因常被沉默;三个编码沉默调控蛋白的基因:SIR2,3,4(SIR:沉默信息调控子),这3个基因所表达的蛋白会形成 SIR2-SIR3-SIR4 复合物;SIR2:去乙酰化酶;下图为酵母端粒的沉默:
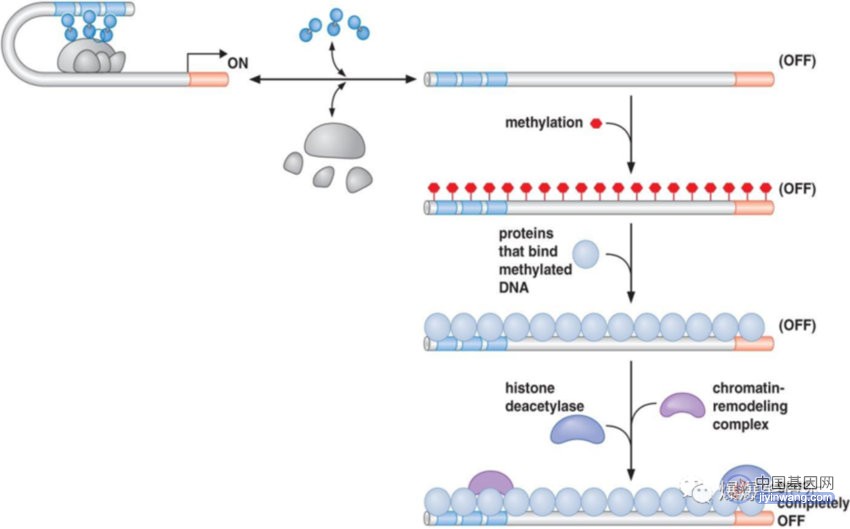
2.2 DNA甲基化的基因沉默
哺乳动物基因组上大段区域被甲基化 DNA 标记,DNA 甲基化通常出现在异染色质区,甲基化 DNA 序列常被招募组蛋白去乙酰化酶和组蛋白甲基化酶的 DNA 结合蛋白(如MeCP2)识别,进而修饰邻近的染色质,因此,DNA 甲基化标记异染色质将要形成的位置;下图为通过DNA甲基化和随后的组蛋白修饰而关闭基因的例子:
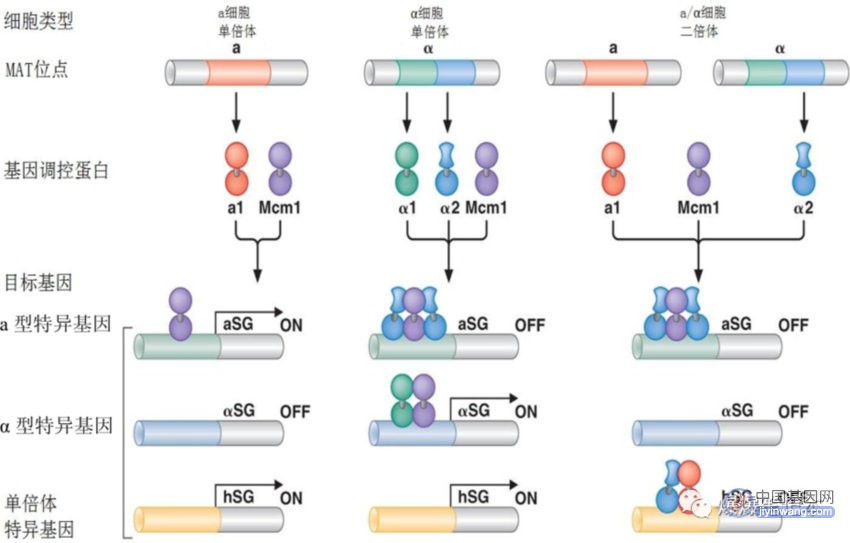
3、啤酒酵母交配型基因的组合调控
组合控制是真核生物复杂性和多样性的核心所在;在真核生物中,存在广泛的组合控制;啤酒酵母细胞以3种形式存在:单倍体交配型a,单倍体交配型α,交配型a和α交配并融合形成的二倍体;a型和α型的不同由其表达两套不同的基因决定;a型特异基因和α型特异基因;由抑制子和活化子的不同组合方式调控;详细机制如下图:
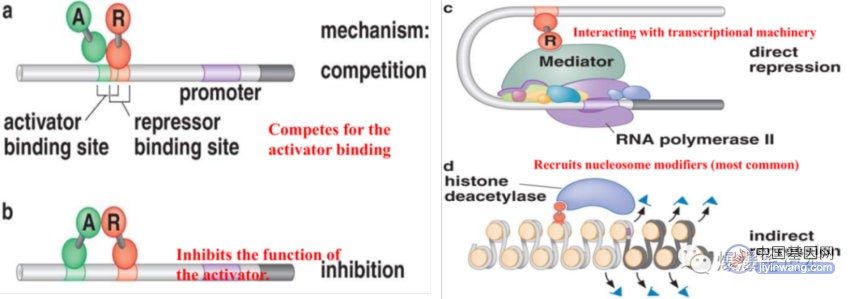
4、真核生物转录抑制子
真核抑制子招募核小体修饰物,使核小体结构更紧密或者去除能够被转录机器识别的基团,例:组蛋白去乙酰化酶去除乙酰基团一些核小体修饰物,在组蛋白尾巴添加上甲基基团,使转录被抑制;真核抑制子的其他作用机制:与活化子竞争重叠结合位点;抑制子与活化子旁边的位点结合,并与活化子相互作用,抑制活化;抑制子与启动子上游位点结合,通过与在启动子处的转录机器作用方式抑制转录起始,如下图:
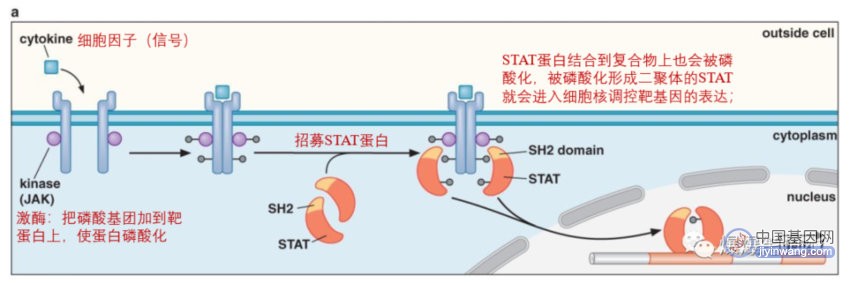
5、信号转导与转录调控蛋白的调控
信号通常通过信号转导通路传到转录调控蛋白;信号指的是起始配体,如糖或蛋白质;在真核细胞中大多数信号通过信号转导通路传到基因,起始配体通常被细胞表面特异受体感应;信号转导通路指的是起始配体(信号)结合细胞表面特异受体的胞外结构域,信号因此传递给该受体的胞内结构域,通过构象改变或者二聚化,信号再被分程传递给相关的转录调控蛋白,转录调控蛋白控制靶基因表达;如STAT通路:
6、基因的表观遗传调控的实例
有些基因表达模式必须得到继承,在发育中一个细胞释放的信号导致邻近细胞开启特异基因,这些基因有时在这些细胞的许多代都保持开启,即使诱导它们开启的信号只是瞬间存在;
表观遗传调控:在缺乏起始信号和缺乏基因突变的情况下,基因表达模式的继承;下图为 λ 噬菌体表观遗传调控的例子:
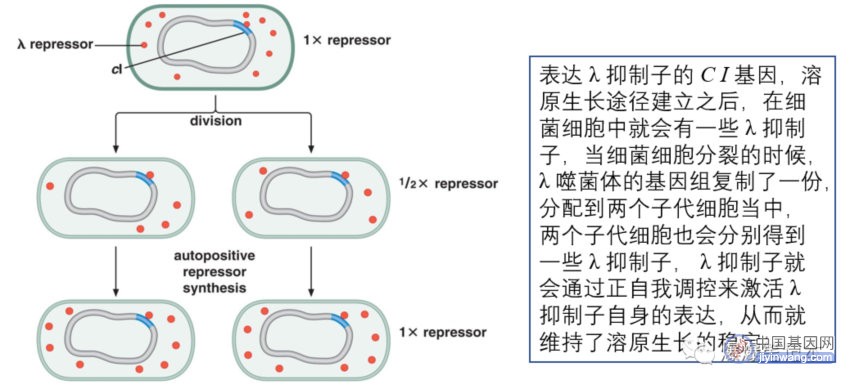
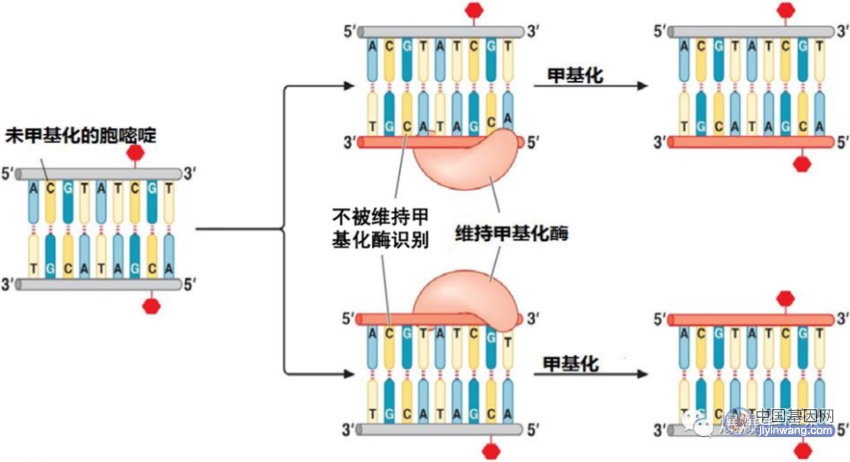
DNA甲基化在细胞分裂中得到保持,也属于表观遗传调控,是通过特定的DNA甲基化酶(维持甲基化酶)的作用来实现的;
声明:本站所有文章资源内容,如无特殊说明或标注,均为采集网络资源。如若本站内容侵犯了原著者的合法权益,可联系本站删除。





")















