抗病基因“假扮”激素受体“诱敌深入”并激活免疫机制
7年前的一个周六晚上,南京农业大学植物保护学院教授陶小荣照例和学生们交流研究进展。学生陈静刚刚做出来的辣椒免疫受体Tsw的电泳图结果引起了他的注意。
“这个受体是同类型其它受体的两倍大小。为什么它会这么大?它到底有什么功能?” 陶小荣觉得,这个看似偶然的现象很可能是揭秘植物抗病机制的重要线索。“只有做重要的科学问题,才能出重大的突破。”
此后,他们在这片“无人区”坚持不懈的探索,并第一次揭示了病原物效应子直接靶向植物激素受体促进侵染,而植物NLR免疫受体模拟受攻击的激素受体监控病毒效应子的干扰,并激活免疫机制的新机制。
12月15日,这一研究成果在线发表于《自然》杂志。中国工程院院士康振生说,这一新的进展,为我国植物免疫学研究揭开了新的一页,提供了新的思路;为抗病育种等应用前景奠定了必不可少的理论基础和技术途径。
一张电泳图引发的研究
电泳图结果中,辣椒免疫受体基因Tsw的大小是6.3kb。而一些经典的NLR免疫受体基因只有3kb。能敏锐地观察到这个“异常”,是由于这个团队已经深耕植物免疫系统研究多年。
陶小荣告诉《中国科学报》,当时他们正在研究番茄斑萎病毒,该病毒是最具破坏性的植物病毒之一,每年在全球范围内对辣椒、番茄等重要经济作物造成了严重的经济损失。

番茄感染番茄斑萎病毒后症状。南京农大供图

辣椒感染番茄斑萎病毒后症状。南京农大供图
然而,经历了千万年的进化,病原微生物和植物之间一直在进行着复杂而精细的进攻与防御的“军备竞赛”。想要在这场纷乱的“战争”中厘清机制,并不是一件容易的事。
陶小荣解释说,病原微生物为了自己的侵染和传播,需要入侵到植物体内,它利用自己的“武器”去攻击寄主植物;而植物为了自身的生存,需要跟病原物去战斗,从而进化出防御系统监控并销毁病原物“武器”的攻击。
“在彼此战斗过程中,病原物的‘武器’会越来越多样化,而植物也会进化出越来越多样化的防御系统,阻止病原物的进攻,这就是植物和病原之间的‘军备竞赛’。”陶小荣说,植物和病原之间的军备竞赛机制是一个复制而精密的系统,搞清楚里面的一条条线索和机制,需要大胆地假设和小心地求证。
“找到重要的线索,这个就是做科研的奥秘所在。”陶小荣说,已知Tsw是辣椒中针对番茄斑萎病毒的抗病基因,它编码一个NLR免疫受体,通过识别病毒编码的效应子NSs蛋白进而诱导植物对病毒的免疫反应。这正是论文共同第一作者、南京农业大学植物保护学院博士生陈静能够在对Tsw的研究中发现重要线索的原因。
陈静介绍,然而Tsw-NLR受体蛋白的一级结构上比对不到什么有用的信息,此前也没有其他相关研究可以参考,无法解析Tsw这么大的功能是什么。因此他们就开始分析该蛋白的三维结构模型,希望找到有用的线索。
“这是一个知识盲区,用现有的知识体系还无法解释这样的现象。前沿研究是无人区。”陶小荣说,这个课题经历过一波三折,克服一道道困难,没有坚持是不可能完成的。
“模拟”激素受体 “诱骗”病毒效应子
通过三维结构建模和同源比对分析发现,在Tsw中具有一个超大的富含亮氨酸重复序列结构域(LRR)。更令人吃惊的是,该结构域竟然与植物激素茉莉酸、生长素和独脚金内酯这三个植物激素的受体相似。
在植物复杂而精细的免疫系统中,由激素激发的免疫途径在抗病过程中发挥重要作用。陶小荣介绍,茉莉酸参与抗病和抗虫,这都是过去众所周知的。生长素参与植物的生长,比如向光性,但也参与对病毒的抗性。独脚金内酯参与植物分蘖,也曾被报道可以抗病和抗线虫。
在激素通路中,激素受体感受到激素后,会启动下游一系列的基因进行转录重编程。茉莉酸受体COI1、生长素受体TIR1和独脚金内酯受体搭档蛋白MAX2都带有一个LRR结构域,该结构域起着启动激素通路的作用。而Tsw-NLR受体蛋白中的LRR结构域正是与激素受体的LRR结构域相似。
“这直接促使我们提出一个‘军备竞赛’的假说:病毒会攻击这三种植物激素的受体,而Tsw这个NLR免疫受体是用来监控病毒的这种攻击的。这是这个课题最重要的科学问题。”陶小荣说。
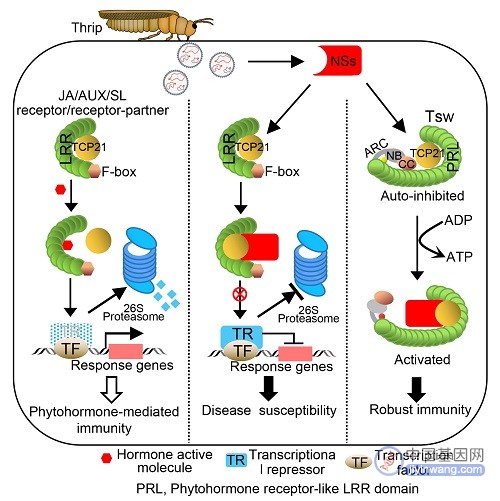
Tsw NLR免疫受体监控病毒靶向激素受体诱导抗病的工作机制图。南京农大供图
过去研究中有不少病原物效应子攻击植物激素通路的报道,但是没有报道过病原物效应子直接攻击植物激素受体的例子,植物免疫系统是否能够监控效应子也不知道。
不过,科学家已经知道,Tsw通过识别病毒效应子NSs,诱导植物产生强烈的抗病反应。“于是我们想到,病毒NSs蛋白会攻击这三种植物激素的受体,而Tsw这个免疫受体则进化出类似于植物激素受体的结构域,通过模拟植物激素受体,‘诱骗’病毒效应子NSs,继而监视病原物攻击过程,最终通过植物免疫系统实现抗病这一过程。”陶小荣说。
“魔”高一尺“道”高一丈
进一步的研究证实了这个猜测。病毒效应子NSs与三种植物激素通路互作时,并不直接与激素受体相互作用,而是与一种名为TCP21的蛋白质直接相结合,TCP21能够与三种激素受体相互作用并抑制激素信号的活性。
陶小荣解释,当TCP21与病毒效应子一结合,就好像形成了一条遏制激素活性的绳索,将激素受体牢牢捆绑在一起,束缚了它们的手脚,活力自然大大降低。这样一来,由激素系统激发的免疫通路就陷入了瘫痪。
激素介导的这一层免疫系统沦陷,植物另一层免疫系统却在暗中储备“兵力”。论文共同第一作者、南京农业大学植物学院博士生赵延晓介绍,植物免疫受体Tsw进化出来模拟植物激素受体的结构域,也与TCP21结合,并巧妙利用了病毒蛋白“捆大绳”的特性,在病毒蛋白NSs存在下,Tsw—NLR免疫受体与TCP21的结合,相对于激素受体与TCP21的结合具有更强的亲和力。
因此,病毒靶向激素受体的时候,NLR免疫受体Tsw会率先感知到这个过程,进而迅速触发免疫反应。
“TCP21同时跟三个激素受体互作,病毒效应子NSs通过靶向TCP21把三种激素受体全部‘干掉’,这是病毒的‘高明’之处。而植物免疫受体Tsw—NLR进化出来模拟植物激素受体的结构域,也与TCP21互作,病毒效应子NSs靶向三个激素的时候,免疫受体Tsw—NLR凭借更强的结合力会提前感知到这个过程,并与病毒效应子NSs结合,激发下游免疫反应。这是植物免疫受体的‘高明’之处。”陶小荣说。
而且,激素介导的抗病是一种比较弱的基础抗病性。NLR免疫受体蛋白介导的抗病性则非常强烈持久,可以有效灭除病原菌。在病毒效应子NSs存在的情况下,植物通过这一机制“选择”了更为强烈的NLR免疫反应,诱敌深入,从而更好的保护自己。“这为作物抗病的生产应用提供了广阔前景。”陶小荣说。
新机制为抗病育种提供新思路
陶小荣强调,没有病原菌时,NLR免疫受体这个“开关”处于关闭状态,这时抗性不启动;当病原菌入侵时,NLR免疫受体感知到效应子,开关就会打开,转换成另外一种状态,启动下游一系列抗病反应。

陶小荣(左)带领学生分析实验现象。南京农大供图
他说,NLR免疫受体也是过去人们所熟知的经典的抗病基因(R基因),是在生产上最有利用价值的抗病基因。这一类的抗病基因的特点是,单基因就可以具有免疫作用,只要将这个抗病基因通过杂交和回交引导感病品种中,感病品种就可以实现抗病性。因此这一类抗病基因在抗病育种中得到广泛应用。
“全球每年因病毒病造成的作物经济损失超过4000亿元。虽然没有有效杀灭病毒病的农药,但是利用抗病品种可以将其有效控制。”陶小荣说。
《自然》审稿专家表示,这一研究结果提供了一个复杂的“效应子毒性—NLR反毒性”的令人感到神奇的案例。这一发现非常精彩,将对植物免疫学相关领域产生深远的影响。
中国科学院遗传与发育生物学研究所研究员周俭民说,病毒是病原微生物中最小最简单的一种,但是人类至今对病毒了解的不多。该研究成果深入研究了抗病毒基因,揭示了前所未知的新的抗病机制,将启发科学家如何影响植物的激素系统,使得植物能够更好的抵抗形形色色的病毒病害。
康振生建议,进一步的研究可以聚焦在如何把新发现的抗病机理与生物育种有机结合起来之上,并为其它作物和病毒病研究提供借鉴和参考。对抗病机制的进一步探索和研究,可以在这些新的知识和理论体系的指导下,发展出新的抗病技术、培育出新的抗病材料。(来源:中国科学报 李晨 许天颖)
相关论文信息:https://doi.org/10.1038/s41586-022-05529-9
声明:本站所有文章资源内容,如无特殊说明或标注,均为采集网络资源。如若本站内容侵犯了原著者的合法权益,可联系本站删除。





")















