探索关于微孢子虫病原体与宿主蚊子的相互作用原理和基因组点位
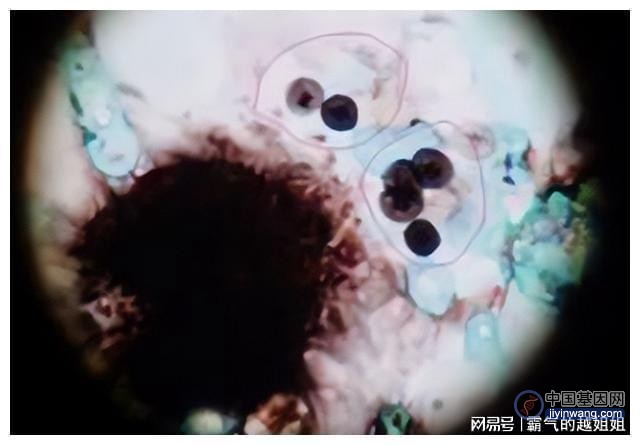
蚊子免疫应答的微孢子虫病原体分析了对伊迪杆菌感染的应答因感染方式的不同而有显著差异,aeedis在感染过程中保持增强的细胞表面修饰和信号能力。
库氏弧菌对宿主的基本代谢物依赖性较低,并保留了剪接体成分的一个子集,其转录组主要集中在生长和复制上。
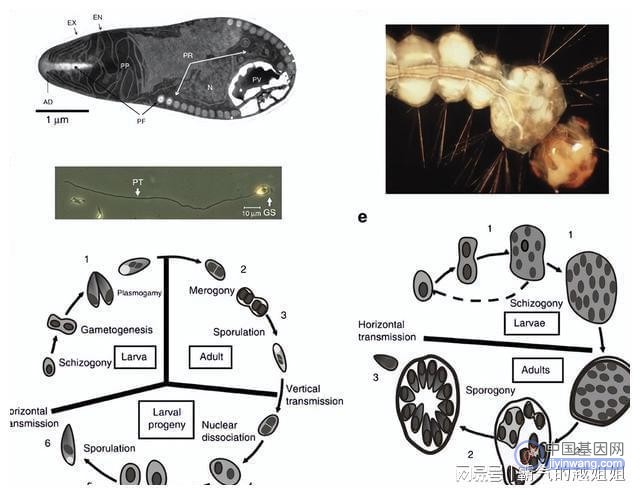
其中专性细胞内病原体依赖于与宿主的复杂相互作用,它们必须在宿主细胞内获取营养和其他代谢物,微孢子虫在孢子中的感染阶段包括一个高度发达的注射装置,用于穿透宿主细胞。
一、剪接蛋白的系统发育分析
环境中的伊迪杆菌孢子被蚊子幼虫摄入后水平传播,聚集在胃盲肠(中肠的一个特殊区域),他们发展成第一个胞内孢子。
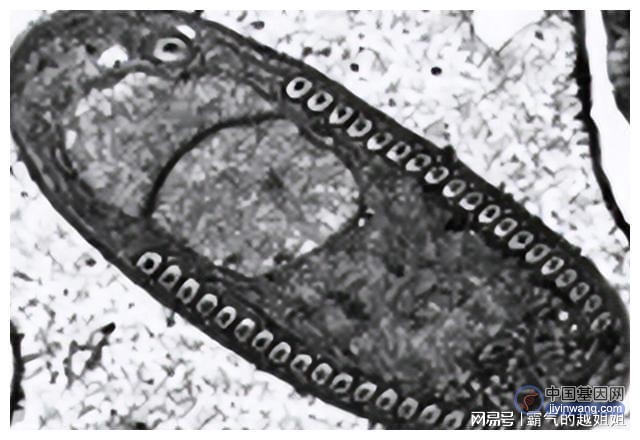
这些孢子穿透墙中肠和入侵宿主细胞称为绛色细胞和血的成年蚊子后发展成第二个胞内孢子和迁移到卵巢。
环境中的孢子被幼虫摄入,穿透中肠壁,并在宿主从幼虫发育到成虫的过程中全身感染宿主,库氏弧菌产生新的孢子并释放到环境。
微孢子虫具有紧凑的基因组,代谢能力降低,基因异常紧密,内含子很少,基因间区很小,微孢子虫已经进化出许多获取这些营养物质的机制,包括用于从宿主细胞“窃取”ATP的ATP转运体。
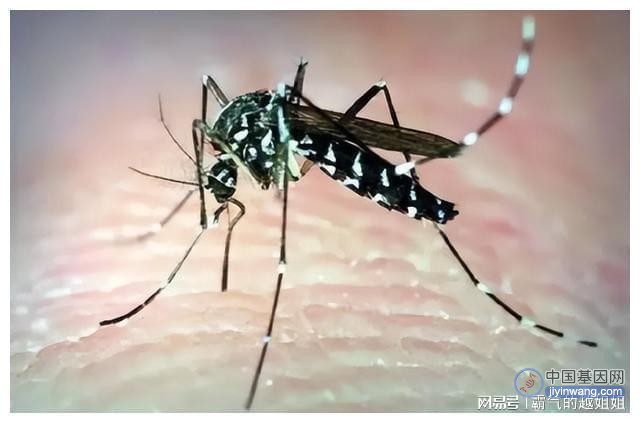
参与宿主-病原体相互作用的其他方面的微孢子虫基因,如逃避宿主免疫反应,分泌的蛋白质可能包括与宿主细胞蛋白相互作用以促进病原体生长的效应物。
尽管基因广泛丢失,但微孢子虫基因组编码了许多扩展的基因家族,特别是具有分泌信号的物种特异性蛋白质,这些蛋白质被假设参与了这些宿主-病原体相互作用。

库氏弧菌基因组每700个碱基含有1个单核苷酸多态性(SNP),平均等位基因平衡为58%,在伊蚊中发现的SNP数量和频率更高,每284个碱基中有1个SNP,平均等位基因平衡为59%。
为了估计这些物种的系统发育位置,我们鉴定了伊蚊、库氏弧菌和另外12种微孢子虫之间的同源物种,aeedis在微孢子虫中拥有最多的蛋白质编码基因反映出aeedis特有的基因。
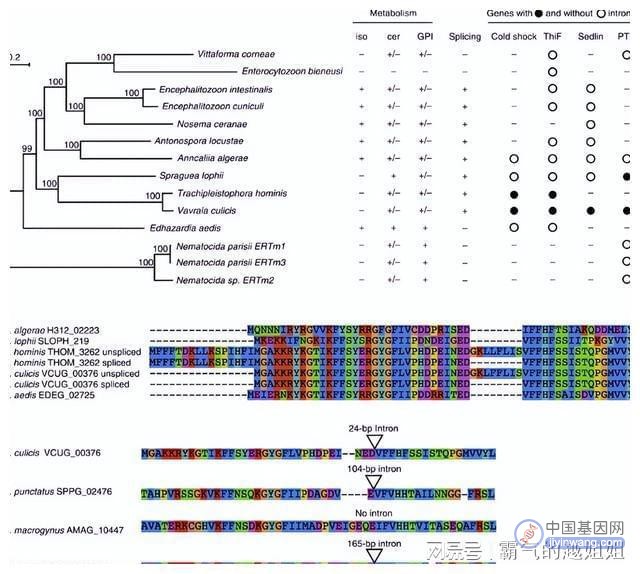
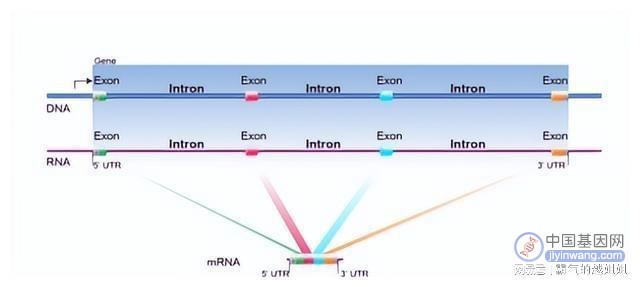
在之前的其他微孢子虫基因组分析中,已经注意到内含子和编码剪接体成分的基因的可变丢失。
为了确定这些微孢子虫生命周期的转录特征,从代表感染主要阶段的时间点对基因表达进行了量化,每个时间点有两个独立的样本。
样本之间表达值的相关性显示,伊迪杆菌转录分为三个主要簇:环境孢子,水平传播的胞内阶段和垂直传播的胞内阶段。
culicis弧菌转录形成两个主要的集群:环境孢子和水平传播的细胞内阶段,没有明显的宿主生命阶段的额外集群。
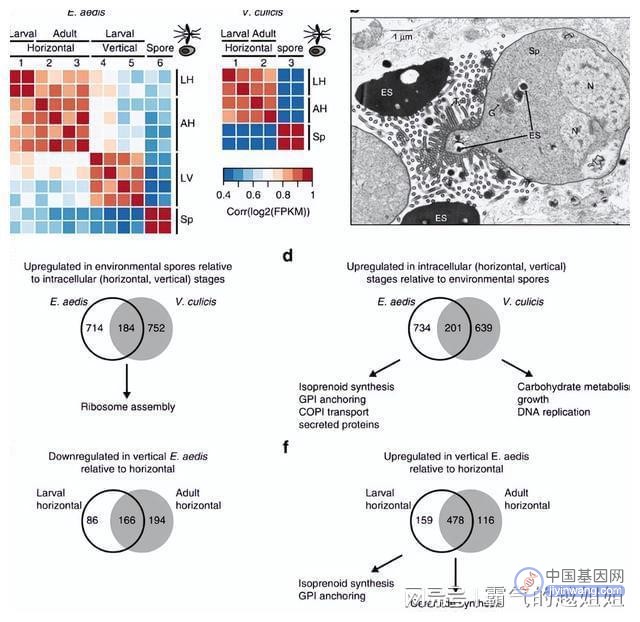
对伊芽孢杆菌和库氏弧菌胞内阶段上调的基因的评估显示,与孢子相比,这两个物种在发育阶段的氧化石墨烯项富集很少重叠。
库氏弧菌中上调的基因在代表生长、碳水化合物代谢和DNA复制的氧化石墨烯项上富集。
aeedis中上调的基因富集于参与蛋白质修饰和运输的途径,如类异戊二烯合成、GPI锚定和参与高尔基-内质网运输的COPI(外壳蛋白)合成。
在aeedis的细胞内阶段,在生命周期的水平或垂直部分特异性上调的基因揭示了代谢潜力的变化以及可以介导宿主相互作用的分泌蛋白的变化。
相比之下,生命周期的水平部分没有显示GO项的富集,比较进食前和进食后的时间点发现,进食后分泌的己糖激酶和海藻化酶均显著上调。
二、aedis性周期中减数分裂基因的表达
考虑到aeedis有性周期明确,而在库氏弧菌中没有观察到减数分裂,我们在转录组时间过程中评估了真菌减数分裂基因的阶段特异性表达。
在一些微孢子虫物种中,HMG两侧的基因,RNA解旋酶和磷酸三糖转运蛋白(TPT)的部分同源性在这些物种中被观察到(但在人猿绦虫中没有明显的同源性)。
每个位点都有两个侧翼基因中的一个,即伊迪杆菌的RNA解旋酶或库利弧菌的TPT,而另一个基因则在每个组合中的不同支架上发现。
所鉴定的9个减数分裂基因在aeedis中至少在一个生命阶段表达,而在V. culicis中存在SPO11和MSH4。

在感染样本中,碱性磷酸酶活性下调,在感染微虫的蜜蜂的肠道中部也观察到同样的效果,中肠入侵过程中,感染样品中的铁稳态和运输上调。
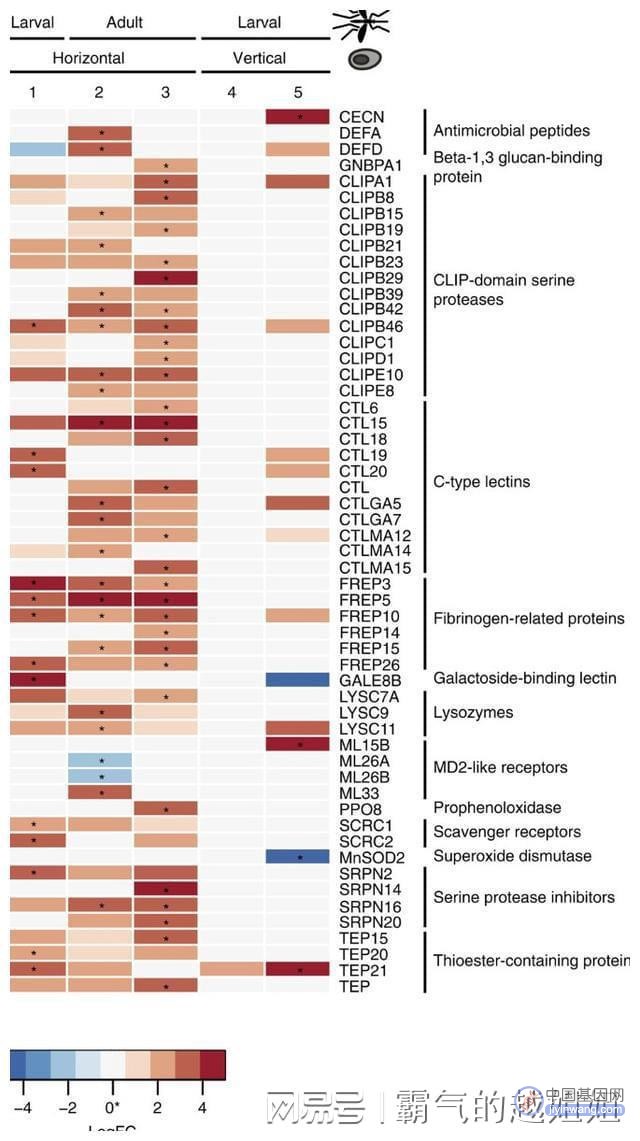
作为纤维蛋白原受体的α -整合素显著上调(AAEL014660, q<0.0001),但不包括在ImmunoDB中。
其他类型的上调基因包括抗菌肽(防御素A和D)、clip结构域丝氨酸蛋白酶、c型凝集素、溶菌酶、丝氨酸蛋白酶抑制剂和含硫酯蛋白,其中一些基因是通过GO项富集分析确定的。
孢子群包含在孢子囊内,并局限于受肌鞘限制的纤维上。
利用ImmunoDB对四斑蝽的基因进行了定位,发现在感染过程中任何时间点(负二项精确检验,q<0.05)唯一差异表达的免疫基因是抗菌防御素(Supplementary Data 7)。
在幼虫期,防御素DEF1在感染过程中显著上调(q<0.05),而在成虫期,DEF1在感染过程中也上调,但不显著(P<0.002, q<0.2),库氏弧菌抑制其蚊子宿主的大部分免疫反应。
对于线虫的变异分析,我们使用GATK unifiedgentyper进行snp鉴定,最佳实践进行筛选(QD<2.0, MQ<40.0, FS>60.0, HaplotypeScore >13.0, MQRankSum< - 12.5, ReadPosRankSum< - 8.0)。
对snp进行筛选,要求GATK基因型质量≥30,读取深度≥10,对于这两个程序集,使用UnifiedGenotyper发布了所有位点的比对统计信息,并用于计算每kb的可调用位置(读取深度至少为10,质量至少为30)的snp估计。
以每ml 1 × 10(3)和1 × 10(4)孢子的剂量,每组100只埃及伊蚊幼虫接触匀浆后的伊蚊孢子24 h,然后每组1000只转入含有3 l去离子水的56 cm × 45 cm × 8 cm塑料培养皿中。
埃及伊蚊(1 × 10(8)和7 × 10(7)孢子的最终产率)提取RNA。
纯化后的aeedis和V. culicis孢子成球,在500 μl的RNA裂解缓冲液中重悬,加入等体积的硅化玻璃微珠(425-600 μm),并使用mini - beadbeat -1 (BioSpec)在48(4,800转/分)条件下机械破坏悬浮液30 s。
细胞计计数证实了孢子的破坏,裂解物从玻璃珠中除去,通过RNA提取处理。
为了证实微孢子虫RNA的成功提取,我们按照基因组DNA的方法对总RNA合成的cDNA进行了qPCR,使用dUTP第二链标记方法为poly(A)选择的RNA样本构建了链特异性文库。
在Illumina HiSeq上对文库进行测序,每个样本平均产生7600万对末端reads (101-nt);所有样本的测序质量得分都很高,平均得分在29.2到34.6之间。
三、基因结构和重复元件的预测
使用Prodigal和Genewise预测了一组初始的蛋白质编码基因,使用Uniref90和可用的微孢子菌蛋白集,需要小于100个氨基酸的蛋白质有良好的支持证据,包括PFAM结构域匹配,非通用基因产物名称分配或两种方法的预测。
从基因集中去除与重复元件(重复元件中发现的匹配蛋白或PFAM结构域或TPSI命中(期望<1e?10和>30%覆盖率)相对应的基因,描述的人类分离物的基因组没有被使用,NCBI中没有可用的组装而不是基因集.
最小和最大内含子大小分别为10和300,具有与基序预测相匹配的拼接读映射的culicis基因被认为具有内含子;没有伊蚊基因与配对的配偶或预测基序拼接读映射。

RNAi机制的存在是通过Argonaute和Dicer的同源物的存在来确定的。
使用Bowtie将RNA-Seq读数与微孢子虫转录本序列对齐,使用RSEM估计转录本丰度,计算每个转录本的tmm标准化FPKM。

四、结论
病原体在寄生周期、宿主免疫应答和寄主适应性方面表现出了多样性和复杂性。 基因家族扩张、基因重组和功能基因的丧失等进化事件体现在该病原体的适应性和致病性方面。
蚊子微孢子虫病原体与其他相关物种之间的基因流动和水平转移现象,这种基因流动对于病原体的遗传多样性和适应性进化.
微孢子虫病原体的相互作用和基因组进化研究对于制定疾病预防和控制策略具有指导意义,也为新药物和疫苗的研发提供了理论基础。
参考文献
[1]李巍. 《肯尼亚狒狒肠道原虫系统进化分析及毕氏肠微孢子虫群体遗传学研究》
[2]黄少康. 《两种微孢子虫的蛋白及对家蚕侵染性的比较研究》
[3]许瑛瑛《蜜蜂孢子虫病的检测与防治研究进展》
[4]杨琼《微孢子虫生物防治的研究进展》
[5]张瑞琳,《微孢子虫及人芽囊原虫混合感染与病原体形态观察》
声明:本站所有文章资源内容,如无特殊说明或标注,均为采集网络资源。如若本站内容侵犯了原著者的合法权益,可联系本站删除。





")















