科学家发现一神秘病毒,没有一个基因与已知生物匹配,仿佛非地球
病毒是一种非细胞生命形态,它由核酸和蛋白质外壳构成,病毒自身缺乏代谢系统,因此病毒离开宿主细胞,就成了没有任何生命活动、也不能独立自我繁殖的化学物质。病毒的分类多种多样,直至今日,仍有新的病毒被不断发现,实际上,人类已经发现的病毒只是全部病毒的冰山一角。近日,巴西米纳斯吉拉斯州联邦大学的PauloV.M.Boratto等人从位于巴西的一个人造城市湖泊中收集浑水样本,从该水样中分理处一种新型感染变形虫的巨型病毒,并将其命名为Yaravirusbrasiliensis(雅拉病毒),雅拉病毒具有令人费解和神秘的起源。雅拉病毒的基因组注释显示,在核苷酸水平上,没有一个基因与已知生物的序列匹配;在氨基酸水平上,仅有6个蛋白与抑制蛋白具有遥远的同源关系。该研究近日以Amysterious80nmamoebaviruswithanear-complete“ORFangenome”challengestheclassificationofDNAviruses为题,发表在生物学预印本网站bioRxiv上。

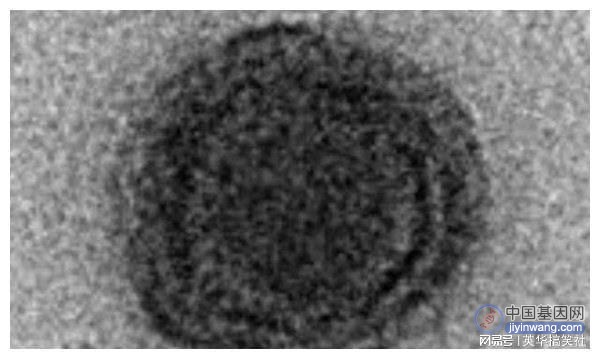
病毒的进化和分类一直是科学界关注和争论的问题,尤其是在发现能够感染原生生物的巨大病毒之后。这些病毒的主要特征是其病毒粒子和基因组的大小很大,它们编码数百至数千种蛋白质,其中很大一部分在目前的公共序列数据库中找不到同源物,这类编码序列通常称为ORFans(孤儿基因),由于缺乏系统发育信息,它们的起源和功能仍然是一个谜。目前所有已知的分离的感染变形虫的巨型病毒均与核质大DNA病毒(NCLDV)有关。该组病毒包括真核病毒家族以及其他巨型病毒谱系。NCDLVs具有双链DNA基因组,根据共享一组祖先垂直遗传基因的标准,该病毒被认是独立起源。雅拉病毒的复制周期雅拉病毒为二十面体,直径约为80nm,纯化病毒颗粒的低温电子显微镜图像表明雅拉病毒具有两个衣壳。感染开始时,雅拉病毒附着在变形虫(阿米巴原虫)质膜的外部,宿主细胞将雅拉病毒内吞到囊泡中,在感染的后期,包含雅拉病毒的囊泡占据细胞核周围的区域,并将其取代,再招募线粒体提供能量,病毒工厂由此产生。
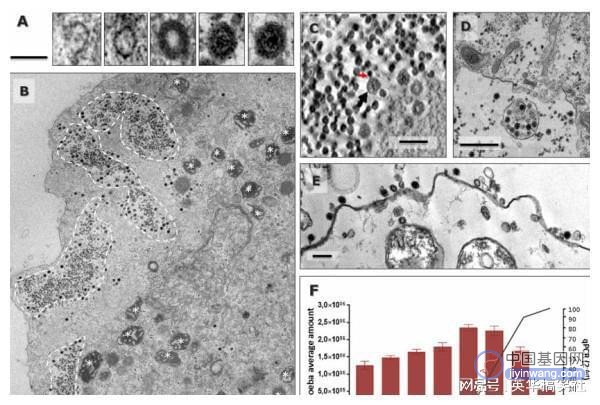
组装方面,病毒复制工厂内显示出现月牙状的结构,再依次添加蛋白质和膜成分,最终形成二十面体结构。雅拉病毒通过胞吐作用和变形虫细胞溶解从而释放子代病毒颗粒。整个复制周期约72小时。
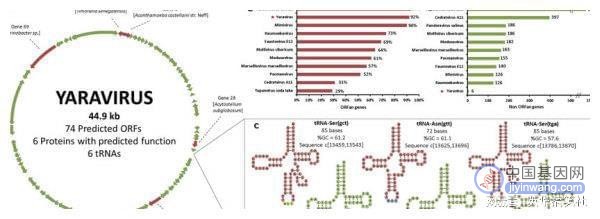
雅拉病毒的基因组经二代测序,雅拉病毒基因组为44924bp的双链DNA分子,并包含总共74个预测基因。尽管基因组比其他巨型病毒小,但雅拉病毒仍编码6个tRNA基因,并均位于基因29和30之间的基因间区域。雅拉病毒基因组的GC含量为46-65%,高于迄今为止发现的任何巨型病毒。
令人惊讶的是,雅拉病毒的基因组注释显示,在核苷酸水平上,没有一个基因与已知生物的序列匹配;在氨基酸水平上,研究者仅发现Pfam-A数据库中的2个蛋白质同源性较高,而nr数据库中的6种蛋白质具有遥远的同源关系。雅拉病毒的蛋白质组从上述资料可知,雅拉病毒的蛋白在公共数据库中没有可检测到的同源物。这促使研究人员更仔细地研究形成雅拉病毒成熟颗粒的蛋白质。蛋白质组学揭示了纯化颗粒中共有26种病毒蛋白,紧接着,研究者使用两个用于域比较的平台Phyre2和Swiss-model,分析了这26种蛋白质的预测三维结构。
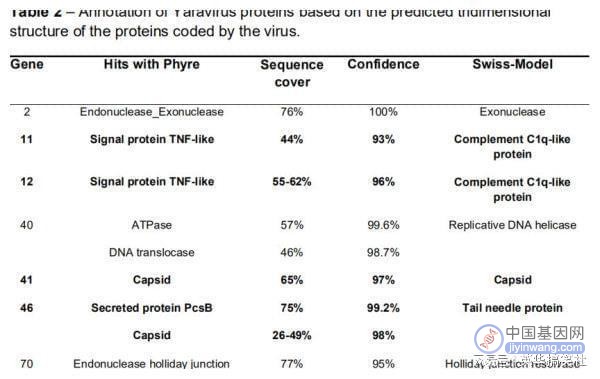
值得注意的是,仅观察到4个序列(基因11、12、41和46)具有与已知蛋白质相似的结构特征,这意味着其病毒颗粒蛋白质组中几乎90%由孤儿基因组成。更有趣的是,其中一种蛋白质与NCLDV主要衣壳蛋白无明显同源性,这可能是新型衣壳蛋白。总而言之,研究者PauloV.M.Boratto等发现了一种新型感染变形虫的巨型病毒——Yaravirus(雅拉病毒)。该病毒在基因组和蛋白质组上与其他巨型病毒相差甚远,极可能是一新的分枝。雅拉病毒与所有其他病毒之间的系统发育距离突显了我们对真核病毒基因组多样性的认知的不足,从而提示我们加强分离新的原生生物病毒研究的必要性。
声明:本站所有文章资源内容,如无特殊说明或标注,均为采集网络资源。如若本站内容侵犯了原著者的合法权益,可联系本站删除。





")















